Arctic biodiversity is structured by a range of environmental factors. To understand which environmental factors that are decisive to the species composition in an area, you need to combine information that vary at different spatial and temporal scales. Within the arctic tundra biome, for instance, the length of the growing season varies between one and four months among different regions and the mean July temperature varies between zero and twelve degrees Celsius. In addition to low air temperatures and short growing seasons the abiotic environmental conditions for life in the Arctic are characterized by low light intensity, which is partly compensated by with midnight sun, by low precipitation, which mainly comes as snow, by strong winds that cause abrasion and desiccation stress, low soil temperatures that affect plant nutrient availability and turnover rates.
Bioclimatic subzones
This large climatic variation is reflected by the vegetation and, accordingly, it is practical to subdivide the Arctic into different bioclimatic sub-zones. Five sub-zones have been recognized, and are referred to as sub-zones A-E with A as the coldest and E the warmest sub-zone (Elvebakk et al. 1999, CAVM Team 2003, Walker et al. 2005) . The subzones A, B and C cover areas that are often referred to as the High Arctic and subzones D and E as the Low Arctic (i.e. sensu Bliss 1997)
The different bioclimatic subzones of the Arctic cover progressively larger areas and increasingly higher habitat and species diversity as one moves from A to E and the vegetation cover and its vertical complexity gradually increases as well. Sub-zone A covers 2% of the non-glacier Arctic with predominately polar deserts, while subzone E has almost continuous vegetation cover dominated by shrub tundra with more than 40 cm tall shrubs.
Arctic land is predominantly treeless permafrost with large areas of tundra. The Arctic environment is highly heterogenous, with for example dry stony ridges, periglacial features, areas of late snow lie, heath, or wet moss, all in close proximity (Thomas et al., 2008). Large areas have recently been reworked by glacial action and possess continuous underlying permafrost, influencing soil hydrology.
On a regional basis, northern areas consist largely of polar desert characterized by low precipitation and a short snow-free growing season. On a landscape scale, the habitat is comprised of a heterogeneous mosaic (Jónsdóttir, 2005). The ridge tops, blown free of winter snow, or areas kept clear of snow by wind eddies, may experience winter temperatures approaching -40°C, while organic soils protected under deeper snow do not exceed temperatures below -10°C (Coulson et al., 1995).
Topography, in combination with snow, strongly modifies the climatic and hydrological conditions experienced by the biota which creates both extensive and small scaled heterogeneity among biological communities within a single subzone. Use guide to explore and compare bioclimatic zones Melting snow and permafrost may provide a constant cold-water source throughout the summer resulting in cold, wet and boggy areas in direct proximity to drier polar desert vegetation. The shallow active layer in the permafrost exaggerates this effect by hindering drainage. Soils may also vary considerably in depth and form between short distances. Generally, the soils are thin, rarely more than a few centimetres deep, and cover moraine debris, patterned ground or bedrock. In wetter areas, moss may develop into thick carpets or turfs some tens of centimetres deep, efficiently insulating the ground beneath against insolation (Coulson et al., 1993).
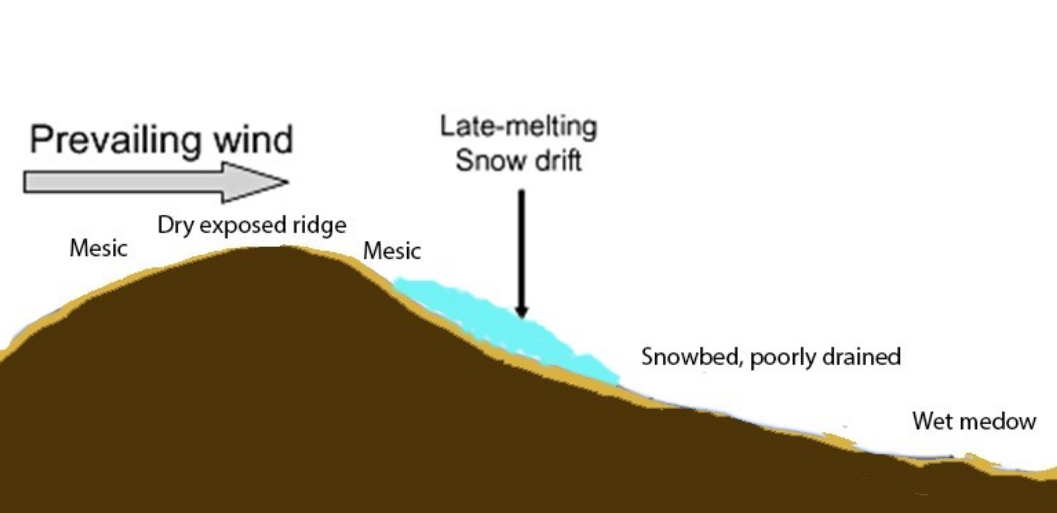
A schematic diagram showing a ridge-snowbed gradient – a topographic gradient that includes a dry exposed ridge, mesic planes/slopes, and a snowbed. A late-melting snowbed will appear above the snowdrift as well and increase habitat diversity. Wet habitats related to the groundwater table will develop in undrained areas.
Adapted by: Pernille B.E.
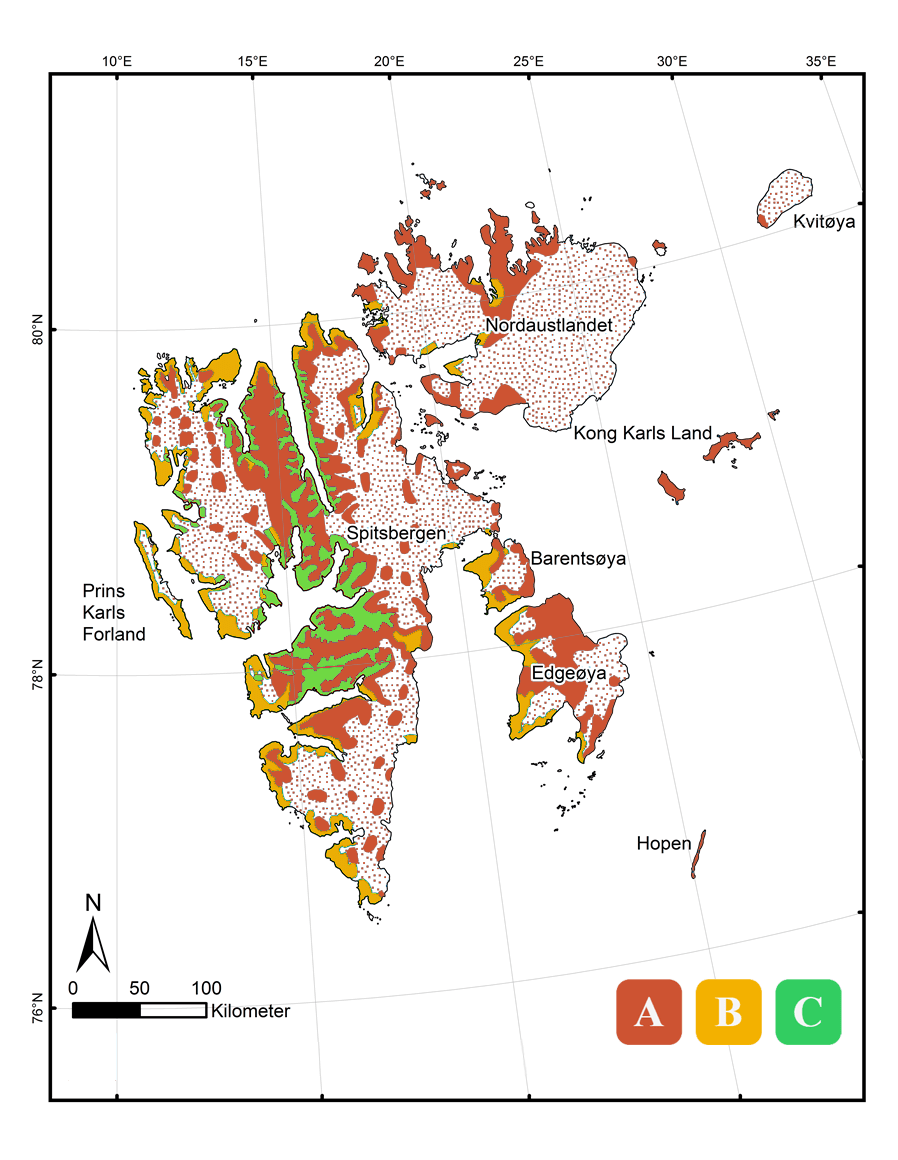
Map showing the three bioclimatic subzones (A, B & C) present in Svalbard.
Source: NTNU Vitenskapsmuseet
- Bliss, L.C. 1997. Arctic Ecosystems of North America. Polar and Alpine Tundra. pp. 551-683.
- Chernov, Yu. I. & Matveyeva, N.V. 1997. Arctic ecosystems in Russia. – In: Wielgolaski, F.E. (Ed) Ecosystems of the World 3. Polar and alpine tundra. Elsevier. p. 361-507.
- Coulson S, Hodkinson ID, Strathdee A, Bale JS, Block W, Worland MR & Webb NR (1993) Simulated climate change: the interaction between vegetation type and microhabitat temperatures at Ny Ålesund, Svalbard. Polar Biology 13: 67-70.
- Coulson, S.J., Hodkinson, I.D., Strathdee, A.T., Block, W., Webb, N.R., Bale, J.S., et al. 1995. Thermal environments of Arctic soil organisms during winter. Arctic and Alpine Research, 27: 365–371.
- CVAM Team 2003 Circumpolar Arctic Vegetaion Map. Scale 1:7,500,000. Conservation of Arctic Flora and Fauna (CAFF) Map No. 1. U.S. Fish and Wildlife Service, Anchorage, Alaska.
- Elvebakk A (1999) Bioclimatic delimitation and subdivision of the Arctic. The species concept in the High North — A Panarctic Flora Initiative,(Nordal I & Razzhivin VY, eds.), p.^pp. 81–112. The Norwegian Academy of Science and Letters, Oslo.
- Elvebakk, A, Elven, R. and Razzhivin, V. Y. 1999 Delimitation, zonal and sectorial subdivision of the Arctic for the Panarctic flora project. In I. Nordal and V.Y. Razzhivin (eds) The species concept in the high north – a panarctic flora initiative. I. Matematisk Naturvitenskaplig Klasse, Skrifter, Ny serie, 38, 375-386.
- Jónsdóttir IS (2005) Terrestrial ecosystems on svalbard: heterogeneity, complexity and fragility from an arctic island perspective. Biology and Environment: Proceedings of the Royal Irish Academy 105B: 155-165.
- Razzhivin, V.Yu., 1999. Zonation of vegetation in the Russian Arctic. – Det Norske Videnskaps-Akademi. I. Mat. Naturv. Klasse, Skrifter, Ny serie 38: 113-130.
- Thomas, D.N., Fogg, G.E., Convey, P., Fritsen, C.H., Gili, J.M., Gradinger, R., Laybourn-Parry, J., Reid, K., Walton, D.W.H. (2008). The Biology of Polar Regions. Oxford University Press, Oxford
- Walker DA, Raynolds MK, Daniels FJA, et al. (2005) The Circumpolar Arctic vegetation map. Journal Of Vegetation Science 16: 267-282.


