During the Last Glacial Maximum (LGM), biodiversity was significantly reduced throughout the Arctic (Willerslev et al., 2014); however, the extent of this biodiversity loss varied. The timing and extent of the LGM ice sheets varied across regions, influencing the dynamics of extinction/colonization, and ultimately shaping the picture of both species- and genetic diversity we see today (Hewitt, 1996).
Genetic diversity
Current species richness is highest for regions that encompassed large refugia during the last glaciation and have maintained land connections to boreal regions after LGM. The current diversity of mammals in low Arctic regions is for instance strongly linked to their relative proximity to boreal habitats. Regions completely covered in ice during the LGM (Dyke 2004, Bennike et al. 2008), but well connected to refugia and to the boreal biome have higher species richness than e.g. Svalbard or Greenland, which lack such direct contact to their likely refugial area.
Richness is further enhanced in regions with extensive mountains where diverse elevations provide additional habitat heterogeneity. Regions with smaller refugia during the last glaciation have much lower species richness, probably because they sustained much fewer species through the last glacial advance.
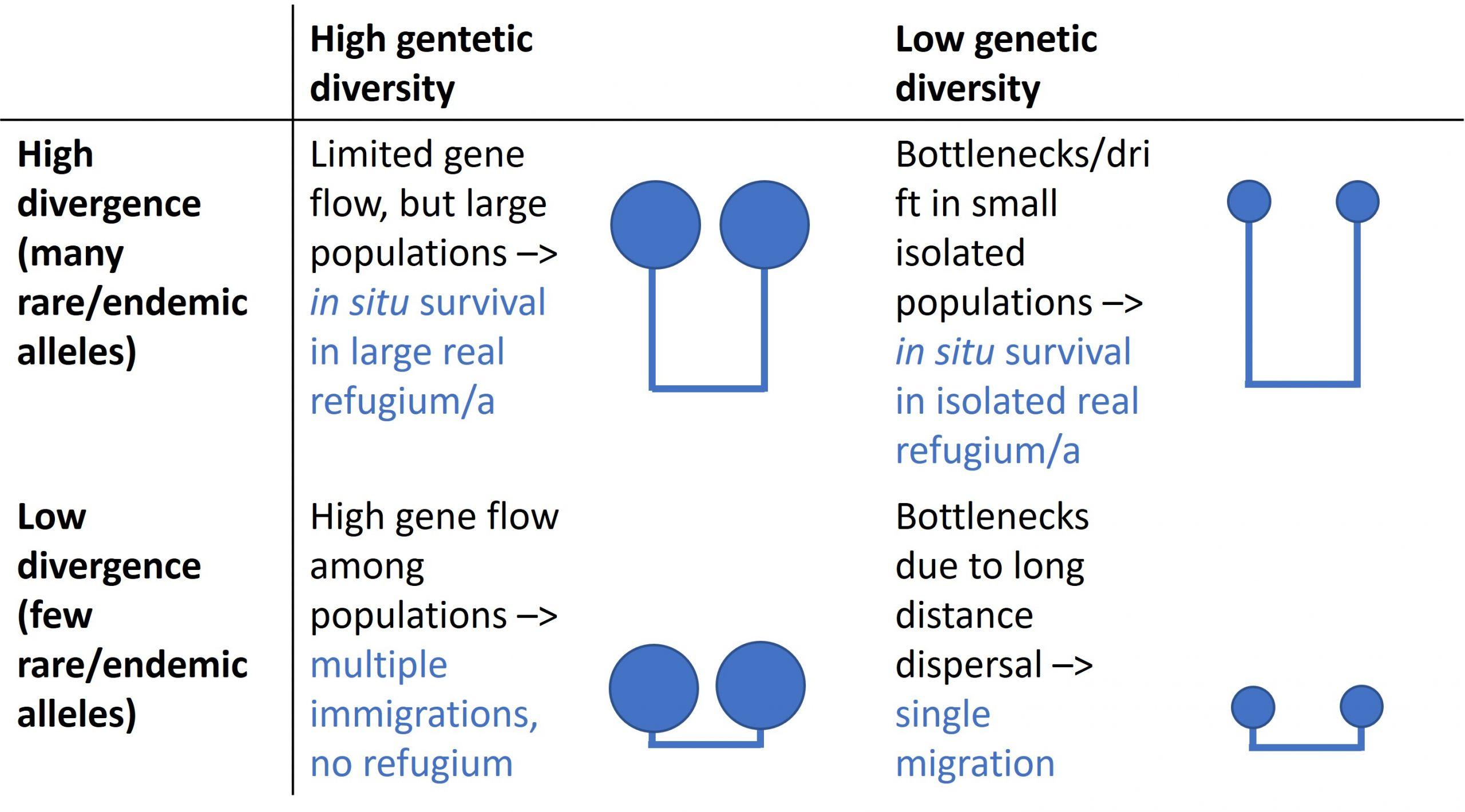
Similar to species diversity, the standard genetic models of glacial refugia and colonization predict lower genetic diversity in formerly glaciated areas and higher genetic diversity in glacial refugia (Hewitt, 1996). The prediction of lower genetic diversity in formerly glaciated areas is founded on the idea that genetic diversity is lost during migration; not all genotypes migrate.
However, this prediction may not hold in areas with secondary contact, where admixture of formerly separated lineages rather increases genetic diversity (Petit et al., 2003). Violation of this prediction is usually most prominent in species with high dispersal capacity. Contrary to traditional beliefs (e.g. Hultén, 1958; Dahl, 1963; Nordhagen, 1963), many plants and animals have been shown to colonize over long distances (de Queiroz, 2005), also in the Arctic (Abbott & Brochmann, 2003).
The prediction of higher genetic diversity within refugia can be fulfilled if the refugia were located within the present distribution range and the population size remained relatively large. This could be the case for the postulated Beringian refugium. The fossil record shows that other larger refugia for arctic organisms were situated south of the ice in Europe and North America. However, these areas are today replaced by boreal or temperate biota (Huntley & Birks, 1983; Tarasov et al., 2000).
Small glacial refugia, such as nunataks protruding above ice caps, are likely to have experienced a drastic reduction in genetic diversity as a result of genetic drift. However, such areas may still contain rare or unique alleles, making them genetically distinct (Schaal et al., 1998, Hewitt, 1999, Stehlik et al., 2001, Hewitt, 2004, Schönswetter et al., 2005). Although the overall importance of glacial survival on nunataks or other small in situ refugia as sources for recolonization after glaciations is debated (Brochmann et al., 2003, Birks et al., 2012, Parducci et al., 2012), there are growing evidence of their existence. Molecular evidence suggests that glacial survival in small in situ refugia was plausible, for example for some plant species, e.g. in inner fjord areas of Eastern Greenland (Westergaard et al. 2011) and in Scandinavia (Westergaard et al, 2019). Lemmings are also suggested to have survived in glacial refugia in Scandinavia (Fedorov & Stenseth, 2001).
Glacial survival in cryptic refugia seems to be more elusive for most invertebrate groups, which to date have pretty much only been defined from large well-known refugia such as the Beringia and the Bank Islands in the Canadian Archipelago. Yet, this is probably most likely linked to the lack of extended research using tools able to identify cryptic refugia than to their absence.
Lectures on refugia and mirgation

Species usually loose genetic diversity during migration.
Source:

Leading edge colonization: (re-)colonization of several species during interglacial periods occurred via long-distance dispersal events, or “leading-edge” colonization, where a few individuals from the source population first colonized and then expanded their range to fill emerging geographic and ecological niches leading to the exclusion of secondary waves of colonizers and thereby reducing genetic diversity at the leading-edge
Source:
Interpretation of phylogenetic patterns using population genetics.
Source:
- Abbott RJ & Brochmann C (2003) History and evolution of the arctic flora: in the footsteps of Eric Hultén. Molecular Ecology 12: 299-313.
- Bennike O, Sørensen M, Fredskild B, Jacobsen BH, Böcher J, Amsinck SL, Jeppesen E, Andreasen C, Christiansen HH & Humlum O (2008) Late Quaternary Environmental and Cultural Changes in the Wollaston Forland Region, Northeast Greenland. Advances in Ecological Research, Vol. 40 p.^pp. 45-79. Academic Press.
- Birks HH, Giesecke T, Hewitt GM, Tzedakis PC, Bakke J & Birks HJ (2012) Comment on “Glacial survival of boreal trees in northern Scandinavia”. Science 338: 742; author reply 742.
- Brochmann C, Gabrielsen TM, Nordal I, Landvik JY & Elven R (2003) Glacial survival or tabula rasa? The history of North Atlantic biota revisited. Taxon 52: 417-450.
- Dahl E (1963) Plant migrations across the North Atlantic Ocean and their importance for the palaeogeography of the region. North Atlantic biota and their history,(Löve Á & Löve D, eds.), p.^pp. 173-194. Pergamon Press, Oxford.
- de Queiroz A (2005) The resurrection of oceanic dispersal in historical biogeography. Trends Ecol Evol 20: 68-73.
- Dyke AS (2004) An outline of North American deglaciation with emphasis on central and northern Canada. Quaternary Glaciations: Extent and Chronology, Vol. Part 2: North America (Ehlers J & Gibbard PL, eds.), p.^pp. 373-424. Elsevier, Amsterdam.
- Fedorov VB & Stenseth NC (2001) Glacial survival of the Norwegian lemming (Lemmus lemmus) in Scandinavia: inference from mitochondrial DNA variation. Proceedings of the Royal Society of London Series B: Biological Sciences 268: 809-814.
- Hewitt GM (1996) Some genetic consequences of ice ages, and their role in divergence and speciation. Biological Journal of the Linnean Society 58: 247-276.
- Hewitt GM (1999) Post-glacial re-colonization of European biota. Biological Journal of the Linnean Society 68: 87-112.
- Hewitt GM (2004) Genetic consequences of climatic oscillations in the Quaternary. Philosophical Transactions of the Royal Society B: Biological Sciences 359: 183-195.
- Hultén E (1958) The amphi-atlantic plants and their phytogeographical connections. Almqvist & Wiksell, Stockholm.
- Huntley B & Birks HJB (1983) An atlas of past and present pollen maps for Europe: 0-13 000 years ago. Cambridge Universty Press, Cambridge.
- Nordhagen R (1963) Recent discoveries in the South Norwegian flora and their significance for the understanding of the history of the Scandinavian mountain flora during and after the last glaciation. North Atlantic biota and their history,(Löve Á & Löve D, eds.), p.^pp. 241-260. Pergamon Press, Oxford.
- Parducci L, Jørgensen T, Tollefsrud MM, et al. (2012) Glacial Survival of Boreal Trees in Northern Scandinavia. Science 335: 1083-1086.
- Petit RJ, Aguinagalde I, de Beaulieu JL, et al. (2003) Glacial refugia: hotspots but not melting pots of genetic diversity. Science 300: 1563-1565.
- Schönswetter P, Stehlik I, Holderegger R & Tribsch A (2005) Molecular evidence for glacial refugia of mountain plants in the European Alps. Molecular Ecology 14: 3547-3555.
- Schaal BA, Hayworth DA, Olsen KM, Rauscher JT & Smith WA (1998) Phylogeographic studies in plants: problems and prospects. Molecular Ecology 7: 465-474.
- Stehlik I, Schneller JJ & Bachmann K (2001) Resistance or emigration: response of the high-alpine plant Eritrichium nanum (L.) Gaudin to the ice age within the Central Alps. Molecular Ecology 10: 357-370.
- Tarasov PE, Volkova VS, Webb T, et al. (2000) Last glacial maximum biomes reconstructed from pollen and plant macrofossil data from northern Eurasia. Journal of Biogeography 27: 609-620.
- Westergaard KB, Alsos IG, Popp M, EngelskjØN T, Flatberg KI & Brochmann C (2011) Glacial survival may matter after all: nunatak signatures in the rare European populations of two west-arctic species. Molecular Ecology 20: 376-393.
- Willerslev E, Davison J, Moora M, et al. (2014) Fifty thousand years of Arctic vegetation and megafaunal diet. Nature 506: 47-51.


